Archives
- 2026-08
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
Performing an ajmaline test in children is problematic for
2019-06-21
Performing an ajmaline test in children is problematic for 2 reasons. First, the test is apparently less sensitive in children than in adults. In fact, in 1 study, a repeat ajmaline challenge performed after puberty unmasked BrS in 23% of relatives with a previously negative drug test performed duri
-
br Conflict of interest br Case presentation
2019-06-21
Conflict of interest Case presentation A 41-year-old man with a history of palpitations refractory to multiple antiarrhythmic agents for several weeks was referred to our department. A 12-lead electrocardiogram (ECG) demonstrated incessant narrow QRS complex tachycardia (cycle length of 580ms)
-
br On Nov every year
2019-06-21
On Nov 25 every year, the International Day for the Elimination of Violence Against Women, and the 16 days of activism that follow it, help to draw the world\'s attention to the scourge of intimate partner violence and other forms of gender-based violence and their harmful effects on women\'s heal
-
Treatment options for elderly AML must be based on four
2019-06-21
Treatment options for elderly AML must be based on four main factors: patient\'s clinical condition, disease characteristics, patient wishes and social support. Thus, the most frail and oldest among the oldest will have the least net benefit from chemotherapy or hematopoietic transplantation, even i
-
We have received funding from the
2019-06-21
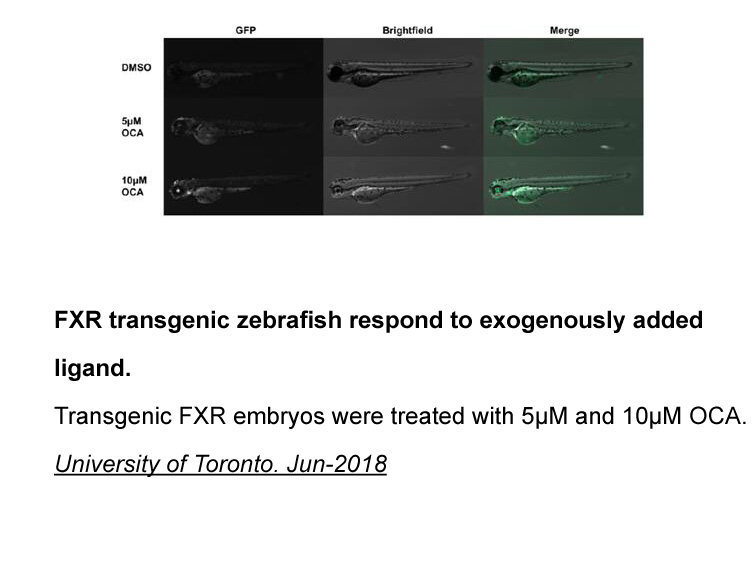
We have received funding from the Wellcome Trust to develop a global atlas of podoconiosis. We aim to advance new knowledge on the geographical distribution and spatial epidemiology of the disease. A first step in this work will be to establish the geographical absence of disease by applying an evid
-
br Material and methods br
2019-06-21
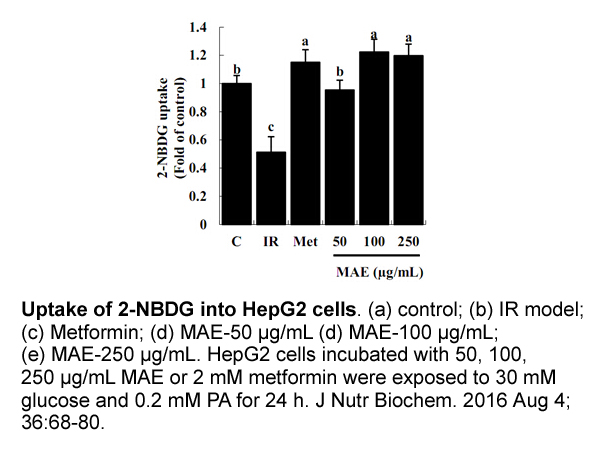
Material and methods Results Discussion ES-SCLC is a disease with poor prognosis. Treatment outcome has not improved significantly during the last decades despite great efforts have been made. Maintenance therapy has been tried in several clinical trials with the aim to prevent the onset of
-
In advanced GIST patients where a course of
2019-06-21
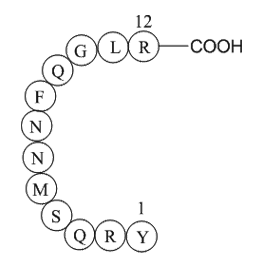
In advanced GIST patients where a course of treatment of Imatinib is interrupted by disease progression, subsequent rechallenge with Imatinib restored tumor control in most patients. In the prospective BFR14 trial, 25 patients with disease progression after interruption of one year of treatment rest
-
Education and lifestyle changes for the prevention
2019-06-21
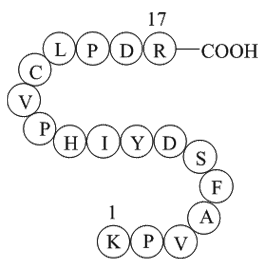
Education and lifestyle changes for the prevention of arrhythmias are critical in BrS. Patients should be informed of the various modulators and precipitating factors that could cause malignant arrhythmias. A prominent S-wave in Lead I has recently been identified as an ECG marker of BrS [298A]. Fev
-
Sin embargo no se puede hablar de
2019-06-21
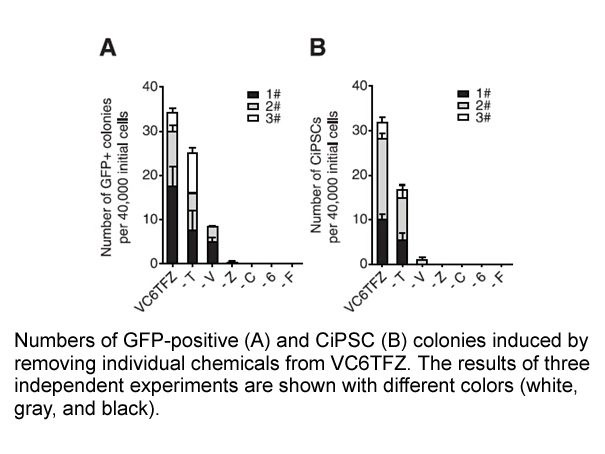
Sin embargo, no se puede hablar de etapas delimitadas con absoluta precisión; la revista Simpleton, en 1998, en un artículo que aparece sin firma, apunta que en la década de los veinte no se ignoraba si ganaría la batalla de la prosa americana el adornado estilo de Fitzgerald o la musculosa dicción
-
In WHO after realising the shortfall in appropriate medical
2019-06-21
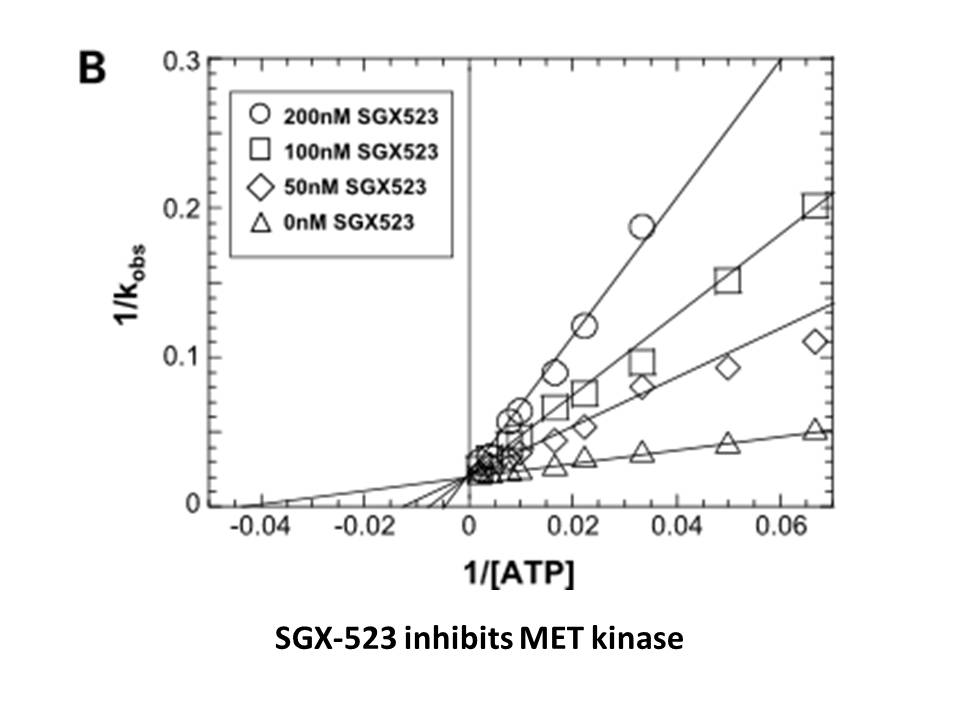
In 2009, WHO, after realising the shortfall in appropriate medical devices in developing countries, called for innovative health technologies to address the technological gap between health-care settings in developed and developing countries. WHO\'s programme, the compendium of innovative health tec
-
Competing pressures for resources across sectors however mea
2019-06-21

Competing pressures for resources across sectors, however, mean that increasing the proportion of resources for health will be difficult. Efficiency gains in the health sector could help to make the case for more investment. Countries need to tackle major sources of inefficiency by doing more of the
-
Beta-Lapachone De acuerdo con esto la mejor par bola es la
2019-06-21
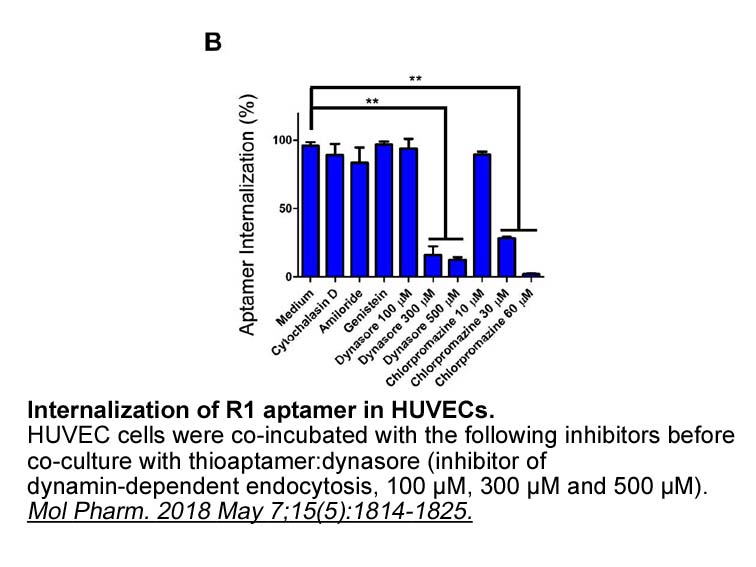
De acuerdo con esto, la mejor parábola es la custodia dorada que contiene un dios vivo y dado Beta-Lapachone ver a los creyentes en la fiesta del Corpus: pura discursividad expuesta y directa que no permite más alegoría que su propia clausura, que no permite más fábula que éste su hacerse en present
-
Las p ginas de este rotativo pusieron
2019-06-21
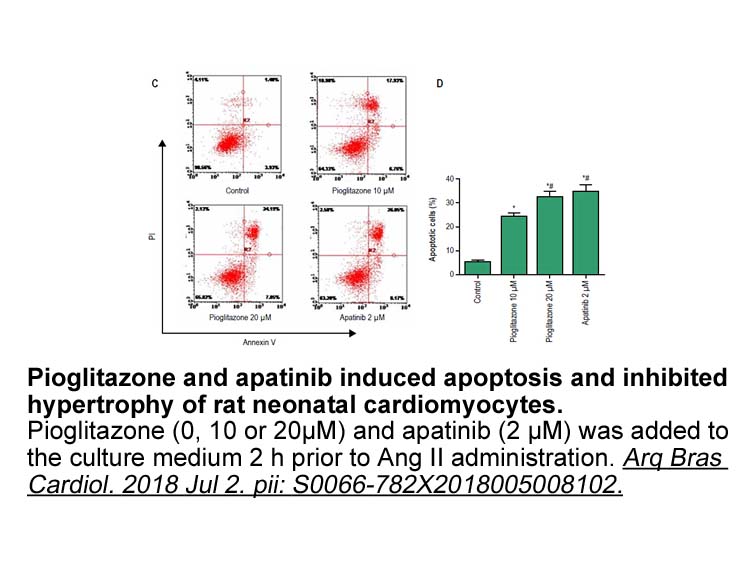
Las páginas de este rotativo pusieron en evidencia las contradicciones y tensiones políticas existentes entre anexionistas e independentistas, La Lucha se mantuvo fiel CW069 su tendencia de crítica a la ocupación militar estadounidense y proclive a evidenciar los efectos negativos de la administraci
-
No evidence has been published from Japan on
2019-06-20
No evidence has been published from Japan on whether or not patients with more than two risk factors have a significantly lower rate of freedom from events than those with less than one risk factor. However, a group from Italy investigated the risk Methylprednisolone Sodium Succinate in individuals
-
br Material and methods The study subjects were consecutive
2019-06-20
Material and methods The study subjects were 28 consecutive patients with organic purchase CGS 35066 disease who underwent ICD implantation for the secondary prevention of sudden cardiac death, based on the indications for ICD implantation in the European Society of Cardiology׳s guidelines [12].
16958 records 1076/1131 page Previous Next First page 上5页 10761077107810791080 下5页 Last page